Aerobic Metabolism
Aerobic Metabolism
Metabolism is defined as the sum of chemical reactions taking place in a live organism to maintain life. Aerobic means oxygen dependent and aerobic metabolism refers to an energy-generating system under the presence of oxygen as opposed to anaerobic, i.e., oxygen independent metabolism. Aerobic metabolism uses oxygen as the final electron acceptor in the electron transport chain and combines with hydrogen to form water [1]. In essence, the vast majority of adenosine triphosphate (ATP) synthesis takes place via aerobic breakdown of energy substrates through the coupling of respiratory chain and oxidative phosphorylation. Aerobic metabolism includes in terms of energy sources carbohydrates and lipids and to a less extent proteins.
Metabolism is defined as the sum of chemical reactions taking place in a live organism to maintain life. Aerobic means oxygen dependent and aerobic metabolism refers to an energy-generating system under the presence of oxygen as opposed to anaerobic, i.e., oxygen independent metabolism. Aerobic metabolism uses oxygen as the final electron acceptor in the electron transport chain and combines with hydrogen to form water [1]. In essence, the vast majority of adenosine triphosphate (ATP) synthesis takes place via aerobic breakdown of energy substrates through the coupling of respiratory chain and oxidative phosphorylation. Aerobic metabolism includes in terms of energy sources carbohydrates and lipids and to a less extent proteins.
Glycolysis
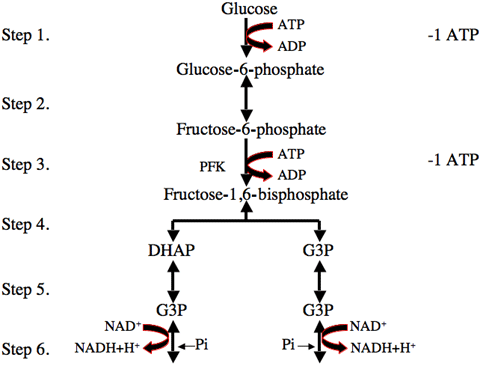
Glycolysis occurs in the cell cytoplasm, and converts one molecule of glucose to two molecules of pyruvate. Two ATP are used in initiating glycolysis but four are generated during the process meaning that two net ATP per glucose molecule are produced during glycolysis. The first ATP donates a phosphate group to glucose to convert it to glucose 6-phosphate and the second is used to add a second phosphate later in the pathway to produce fructose 1,6-bisphosphate. In representations of this pathway aldolase is usually shown (as in the diagram below) to convert one fructose 1,6-bisphosphate molecule to two glyceraldehyde 3-phosphate molecules. In fact it splits it into one glyceraldehyde 3-phosphate molecule and one dihexoacetone phosphate but these are rapidly interconverted by triose phosphate isomerise.
Following glycolysis, pyruvate may be converted into acetyl coenzyme A (acetyl CoA) and continue in the aerobic respiration pathway through the citric acid cycle. Alternatively it may be converted to lactate or used to synthesise fatty acid or ketone bodies. The former occurs if oxygen levels are low, producing lactate for use in anaerobic respiration. The latter occurs when excess ATP is present in the cell, and the rate of the citric acid cycle slows down. As less acetyl CoA is used in the cycle it accumulates and the excess it converted to ketone bodies and fatty acids for eventual storage.
Citric Acid Cycle
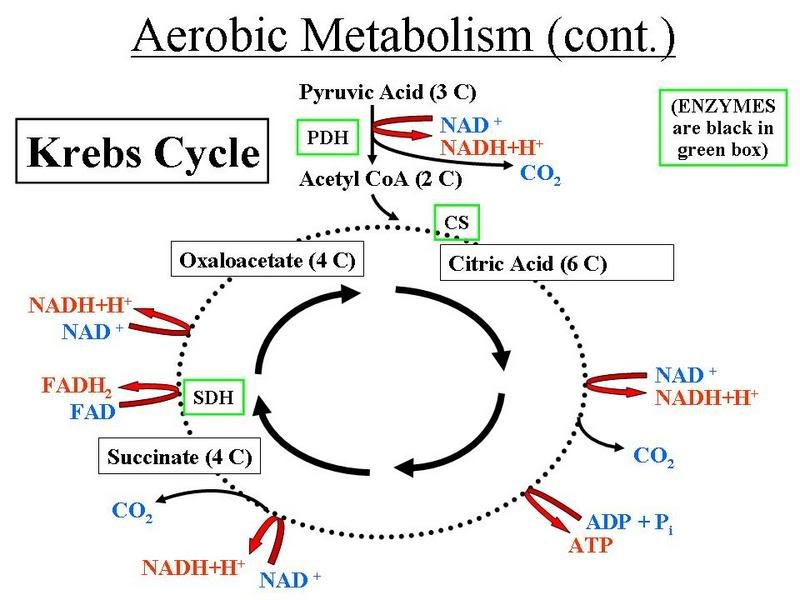
The citric acid cycle (sometimes called the Krebs cycle) is the second phase in aerobic respiration. It occurs in the mitochondria of eukaryotes and its input molecule is acetyl CoA which comes mainly from glycolysis but also from the breakdown of fatty acids and other sources. Acetyl CoA enters the cycle by transferring the acetyl group to oxaloacetate to form citrate. By the end of the cycle the oxaloacetate is regenerated and acetyl CoA is broken down into CoA, CO2 and water.
The citric acid cycle produces intermediate products which can be removed from the cycle and converted to other useful biomolecules. Examples include α-ketoglutarate which is a precursor for amino acid and nucleotide synthesis and oxaloacetate which is a substrate for gluconeogenesis. They also function in regulating the cycle in a negative feedback mechanism where citrate inhibits the incorporation of new acetyl CoA into the cycle and succinyl CoA inhibits its own formation.
Introduction
Respiration is the process by which cells use energy released from breaking covalent bonds to convert adenosine diphosphate (ADP) to adenosine triphosphate (ATP) which functions as the immediate energy source for most metabolic processes. The energy stored in ATP is released by ATPases which break the bond connecting the third phosphate group. Some enzymes bind directly to ATP such as kinases which use it as a phosphate donor. The two forms of respiration are aerobic respiration which requires oxygen and anaerobic respiration which does not. Anaerobic respiration is less energetically efficient than aerobic respiration and is generally used by organisms adapted to a low oxygen environment or as a last resort when oxygen is unavailable.
Aerobic Respiration
Aerobic respiration breaks down into three stages; glycolysis, the citric acid cycle (or Krebs cycle) and the electron transport chain. The first, glycolysis, occurs in the cytoplasm and converts glucose and other sugars into pyruvate. This is then converted to acetyl coenzyme A and oxidised in the citric acid cycle in the mitochondria. Both glycolysis and the citric acid cycle contribute to the production of ATP. The electron transport chain transfers electrons from a donor (usually NADH) to an acceptor, (usually oxygen) though a series of carriers in the inner mitochondrial membrane generating a proton gradient. The energy stored in the form of the gradient is released through ATP synthase to drive ATP synthesis.
Glycolysis
Glycolysis occurs in the cell cytoplasm, and converts one molecule of glucose to two molecules of pyruvate. Two ATP are used in initiating glycolysis but four are generated during the process meaning that two net ATP per glucose molecule are produced during glycolysis. The first ATP donates a phosphate group to glucose to convert it to glucose 6-phosphate and the second is used to add a second phosphate later in the pathway to produce fructose 1,6-bisphosphate. In representations of this pathway aldolase is usually shown (as in the diagram below) to convert one fructose 1,6-bisphosphate molecule to two glyceraldehyde 3-phosphate molecules. In fact it splits it into one glyceraldehyde 3-phosphate molecule and one dihexoacetone phosphate but these are rapidly interconverted by triose phosphate isomerise.
Following glycolysis, pyruvate may be converted into acetyl coenzyme A (acetyl CoA) and continue in the aerobic respiration pathway through the citric acid cycle. Alternatively it may be converted to lactate or used to synthesise fatty acid or ketone bodies. The former occurs if oxygen levels are low, producing lactate for use in anaerobic respiration. The latter occurs when excess ATP is present in the cell, and the rate of the citric acid cycle slows down. As less acetyl CoA is used in the cycle it accumulates and the excess it converted to ketone bodies and fatty acids for eventual storage.
Citric Acid Cycle
The citric acid cycle (sometimes called the Krebs cycle) is the second phase in aerobic respiration. It occurs in the mitochondria of eukaryotes and its input molecule is acetyl CoA which comes mainly from glycolysis but also from the breakdown of fatty acids and other sources. Acetyl CoA enters the cycle by transferring the acetyl group to oxaloacetate to form citrate. By the end of the cycle the oxaloacetate is regenerated and acetyl CoA is broken down into CoA, CO2 and water.
The citric acid cycle produces intermediate products which can be removed from the cycle and converted to other useful biomolecules. Examples include α-ketoglutarate which is a precursor for amino acid and nucleotide synthesis and oxaloacetate which is a substrate for gluconeogenesis. They also function in regulating the cycle in a negative feedback mechanism where citrate inhibits the incorporation of new acetyl CoA into the cycle and succinyl CoA inhibits its own formation.
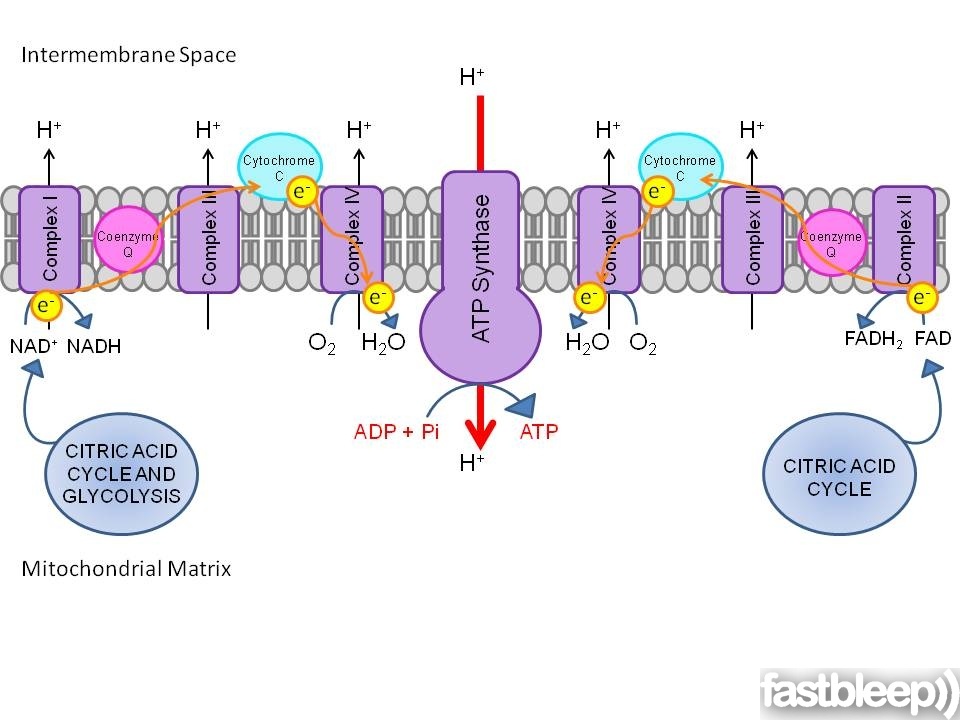
The Electron Transport Chain
The respiratory electron transport chain occurs in the mitochondria of eukaryotes. Electrons are released from a donor molecule and passed along a series of carriers to a final acceptor molecule releasing energy in each step. This energy is used to transport protons (H+) from the inner mitochondrial matrix to the intermembrane space creating a electrochemical and a pH gradient. It is energetically favourable for the protons to cross back into the mitochondrial matrix. The energy released when this happens is used by the enzyme ATP synthase (Complex V) to convert ADP + Pi to ATP. The catalytic part of ATP synthase is coupled to an ion pore, and the energy released as the electrons flow down the electrochemical gradient is used to drive the reaction.
Glycolysis and the citric acid cycle reduce NAD+ to NADH, making NADH a strong reducing agent or electron donor. Oxygen is a strong oxidising agent or electron acceptor. The strong reduction potential of oxygen is the reason why, when available, it is usually the electron acceptor of choice. Anaerobic organisms use other molecules with lower reduction potentials as the electron acceptor meaning that less energy is released per acceptor. By the end of the reaction NAD+ is reformed for reuse and oxygen is reduced to water.
The electron carriers consist of four complexes. Complex I (or NADH dehydrogenase) removes two electrons from NADH and transports four protons into the intermembrane space. The two electrons are then transferred to Coenzyme Q, a lipid-soluble molecule which floats freely in the membrane. Complex II (succinate dehydrogenase) provides an alternative source of electrons for Coenzyme Q from the citric acid cycle producuct FADH. Complex III (Cytochrome c-Coenzyme Q oxidoreductase) transfers the electrons from Coenzyme Q to two molecules of cytochrome c. Cytochrome c is water soluble and exists in the intermembrane space. This process translocates another six protons. Complex IV (cytochrome c oxidase) transfers four electrons from four cytochrome c molecules to oxygen molecules. The resulting negatively charged oxygen atoms bind to free protons to form water.